DNA Eşlenmesi (Replikasyon)
DNA molekülü, her hücre bölünmesi öncesinde kendisini eşleyerek kalıtsal bilginin yavru hücrelere eşit miktarda dağılımını sağlar. DNA eşlenmesinin nasıl gerçekleştiğini ispatlamak için 1958 yılında Matthew Meselson (Methiv Meselsın, 1930- ) ve Franklin Stahl (Franklin Sıtal, 1929- ) tarafından deneyler yapılmış ve farklı modeller test edilmiştir. Meselson ve Stahl yaptıkları çalışmalar ile DNA molekülündeki her iki ipliğinde kendisini eşleyerek yeni birer iplik oluşturduğunu gözlemlemiş ve “yarı korunumlu (semikonservatif) eşlenme” modelinin doğruluğu ispatlanmıştır.
Meselson ve Stahl, yaptıkları deneyde bakterilerin gelişme ortamlarına izotop azot atomunu eklediler.
BİLGİ :
İzotop: Atom veya proton numaraları aynı kütle numaraları farklı elementlere denir. Örneğin 14N ve 15N birbirinin izotopudur.
BİLGİ : Bir DNA molekülünün her iki ipliğindeki azotların kütle numaraları 14 olursa yani (14N14N) şeklinde ise bu DNA molekülü normal azotlu DNA olarak adlandırılır. DNA molekülünün her iki ipliğindeki azotların kütle numaraları 15 olursa yani (15N15N) şeklinde ise bu DNA molekülü ağır azotlu DNA olarak adlandırılır. Eğer DNA molekülünün bir ipliğindeki azotun kütle numarası 14 diğer ipliğindeki azotun kütle numarası 15 olursa yani (14N15N) şeklinde ise bu DNA molekülü melez DNA olarak adlandırılır. Bu DNA’ların , santrifüjleri sonucu deney tüpünde farklı batnlaşmalar görülür. 15N15N formuna sahip ağır DNA molekülleri dibe çökerken , 14N15N formundaki melez DNA molekülleri ortada, 14N14N formundaki normal azotlu DNA molekülleri ise en üstte bantlaşma gösterirler.

Meselson ve Stahl, tarafından yapılan deneylerin aşamaları aşağıda özetlenmiştir:
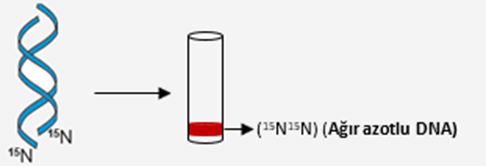
A ) Meselson ve Stahl çalışmalarında E. coli bakterilerini kullanarak azotun ağır izotopu olan 15N içeren kültür ortamında birçok nesil verecek şekilde üremelerini sağlamışlardır. Nesiller sonra ortamdaki bakteri DNA’larının 15N izotopunu taşıdığı gözlemlenmiştir. Bu bakterilerin DNA’ları ayrıştırılıp santrifüj edildiğinde DNA’ların tüpün en alt kısmında bir bant oluşturacak şekilde toplandıkları görülmüştür.
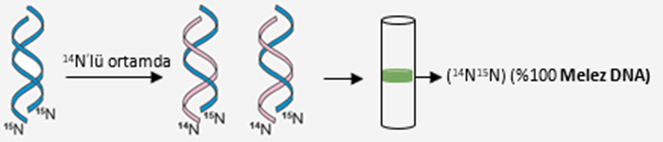
B ) DNA’larında 15N bulunan E.coli bakterileri , 14N izotoplu azotun bulunduğu kültür ortamına bırakılmıştır. Birinci üreme sonucunda bakteri DNA’ları ayrıştırılıp santrifüj edildiğinde deney tüpünün orta kısmında bir bantlaşma olduğu gözlemlenmiştir. Orta kısımda bantlaşmanın nedeni birinci bölünme sonucu meydana gelen bakteri DNA’larının %50 15N, %50 14N (%100 melez DNA) taşımasıdır.
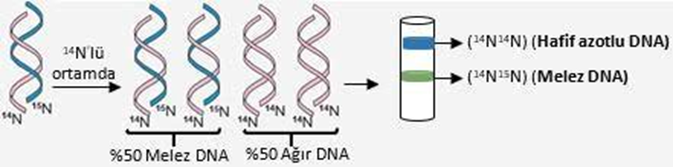
C ) İkinci üreme sonunda santrifüj edildiklerinde ise oluşan bakteri DNA’larının %50’sinin melez (14N15N) olduğu, %50’sinin normal azot (14N14N) içerdiği, bu nedenle de hem ortada hem üstte bantlaşma olduğu gözlemlenmiştir.
SONUÇ : Çalışmaların sonucunda DNA’nın bir ipliği her eşlenme sonucu kendisini aynen korunurken diğer ipliğin ortamdaki izotop azot atomlarını kullanarak sentezlendiği gözlemlenmiştir. Bunun sonucunda DNA molekülü kendisini “yarı korunumlu eşler” olarak ifade edilmiştir.
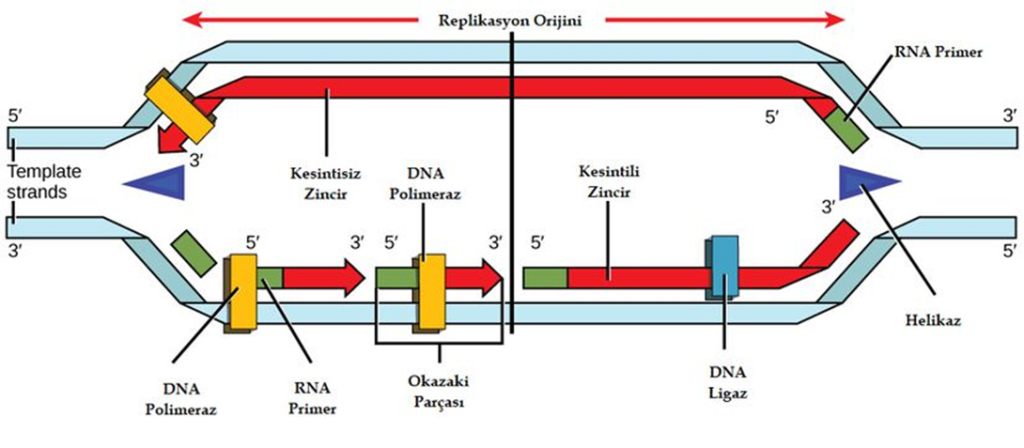
DNA’NIN EŞLENME MEKANİZMASI
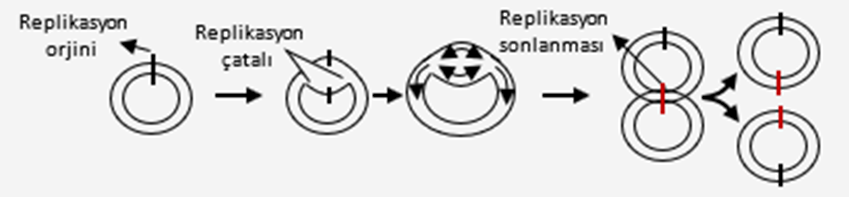
DNA eşlenmesinin başlayacağı özel bölgelere replikasyon orjini adı verilir. Prokaryotlarda halkasal DNA da bir başlangıç ve bir bitiş noktası, ökaryotlarda ise doğrusal DNA üzerinde çok sayıda başlangıç ve bitiş noktası bulunur.
a-) Prokaryotlarda DNA Replikasyonu :
Prokaryot canlılarda halkasal yapıda DNA bulunduğu için bir noktadan başlayan replikasyon , zıt yönde ve her iki iplikte aynı anda devam ederek DNA tamamen kopyalanıncaya kadar sürer.
BİLGİ : DNA replikasyonunu sitoplazmada gerçekleştiren bir canlı kesinlikle prokaryottur.
b-) Ökaryotlarda DNA Replikasyonu :
Ökaryot canlılarda DNA doğrusal ve çok daha uzundur. Ökaryot DNA’larında replikasyon sırasında yüzlerce hatta binlerce replikasyon orjini oluşur. Bu sayede DNA eşlenmesi daha kısa sürede tamamlanır.
Prokaryotlarda olduğu gibi ökaryotlarda da DNA replikasyonu her replikasyon orjininden her iki yöne doğru ilerler , replikasyon kabarcıkları oluşur , her replikasyon kabarcığının iki ucunda replikasyon çatalı vardır. DNA eşlenmesi sırasında helikaz enzimi , çift sarmal ipliklerin replikasyon orjin bölgelerine gelerek iki kolu tersine büküp açar,yeni DNA ipliklerinin sentezinin başlayabilmesi için gerekli olan RNA primerleri ise primaz enzimi tarafından sentezlenir , primaz enzimi bu sayede yeni DNA ipliğinin yapımı için başlangıç noktasını göstermiş olur. DNA polimeraz enzimi, ayrılmış olan her DNA ipliğinin karşısına uygun yeni nükleotitleri sıralayarak kalıp DNA ipliklerine uygun birer DNA ipliği daha oluşmasını sağlar. Sentezlenen yeni iplikler farklı şekillerde uzar. DNA polimeraz enzimi, kalıp ipliğin sadece 3´ ucundaki nükleotidin karşısına yeni nükleotit ekleyebilir. Bu yüzden DNA replikasyonu, 5’ → 3’ yönünde ilerler; Kalıp olarak görev gören zincir 3’ → 5’ yönünde okunur. 3´→ 5´ yönünde uzanan kalıp ipliğin karşısına gelecek olan yeni iplik, 5´ ucundan 3´ ucuna doğru kesintisiz olarak sentezlenir. Diğer kalıp DNA ipliği ise 5´→3´ yönünde uzanır , karşısında oluşacak yeni iplik helikaz enziminin açtığı bölgeye ters yönde uzar. Bu nedenle bu ipliğin kesintisiz bir şekilde sentezlenmesi mümkün olmaz. Replikasyon çatalı açıldıkça yeni iplik, 5´ ucundan 3´ ucuna doğru uzanan parçalar hâlinde sentezlenir. Bu parçalara Okazaki parçaları (fragmentleri) adı verilmiştir. Oluşan bu parçaların bir süre sonra DNA ligaz enzimi ile birbirine bağlanmasıyla kesintisiz bir iplik elde edilir. Böylece her iki iplikte eksiksiz sentezlenmiş olur.